
Kaktüsgiller
| Kaktüs | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||
| Biyolojik sınıflandırma | |||||||||||
| |||||||||||
| Altfamilyalar | |||||||||||
 | |||||||||||
| Dağılımı | |||||||||||
| Sinonimler | |||||||||||
| |||||||||||


Kaktüs Caryophyllales takımının[3] 1750 kadar bilinen türüne ve yaklaşık 127 cinsine sahip Cactaceae bitki familyasının bir üyesidir. "Kaktüs" kelimesi Latince aracılığıyla, Theophrastus tarafından kimliği kesin olmayan dikenli bir bitki için kullanılan Antik Yunanca (yunanca:κάκτος, okunuşu:kaktos) kelimesinden türemiştir.[4]
Kaktüsler çok çeşitli şekil ve boyutlardadır. Çoğu kaktüs, en azından biraz kuraklığa maruz kalan habitatlarda yaşar. Birçoğu, dünyanın en kurak yerlerinden biri olan Atacama Çölü 'nde bulunsa bile, aşırı derecede kuru ortamlarda yaşarlar. Kaktüsler suyu korumak için birçok uyum gösterirler. Hemen hemen tüm kaktüsler sukulent dir, yani kalınlaşmış, etli kısımları suyu depolamak için uyarlanmıştır. Diğer birçok sulu meyvenin aksine, gövde, bu hayati sürecin gerçekleştiği çoğu kaktüsün tek parçasıdır. Çoğu kaktüs türü gerçek yapraklarını kaybetmiş, yalnızca oldukça değişmiş yapraklar olan dikenler kalmıştır. Dikenler, otoburlara karşı savunmanın yanı sıra, kaktüse yakın hava akışını azaltarak ve biraz gölge sağlayarak su kaybını önlemeye yardımcı olur. Yaprakların yokluğunda, büyümüş gövdesi fotosentez yapar. Kaktüsler Amerika'nın güneyindeki Patagonya 'dan kuzeyindeki batı Kanada 'ya olmak üzere - Afrika'da ve Sri Lanka'da yetişen Rhipsalis baccifera haricinde genelde Amerika kıtasına özgüdür.
Kaktüs dikenleri areole denilen, oldukça küçültülmüş bir dal türü olan özel yapılardan üretilir. Areoller, kaktüslerin belirleyici bir özelliğidir. Dikenlerin yanı sıra, areoller genellikle boru şeklinde ve çok boyutlu çiçek açarlar. Pek çok kaktüsün kısa büyüme mevsimi ve uzun uyku dönemleri vardır ve yer yüzeyine ulaşan suyu hızla emen geniş ama nispeten sığ bir kök sisteminin yardımıyla herhangi bir yağmura hızlı tepki verebilir. Kaktüs gövdeleri genellikle nervürlü veya yivlidir bu da yağmurdan sonra hızlı su emilimi için kolayca genişlemelerine ve büzülmelerine ve ardından uzun kuraklık sürelerine imkan verir. Diğer sukulent bitkiler gibi kaktüslerin çoğu da fotosentezin bir parçası olarak "crassulacean asit metabolizması" (CAM) adı verilen özel bir mekanizmayı kullanır. Karbondioksitin bitkiye girdiği ve suyun çıktığı terleme, fotosentezle aynı anda gündüzün değil geceleyin olur. Bitki aldığı karbondioksidi malik asit olarak depolar, onu gün ışığı dönene kadar tutar ve ancak o zaman fotosentezde kullanır. Terleme daha soğuk, daha nemli gece saatlerinde gerçekleştiği için su kaybı çok azalır.
Daha küçük kaktüslerin çoğu suyu depolamak için mümkün olan en büyük hacmi ve terleme kaynaklı su kaybını azaltmak için de olası en az yüzey alanını birleştiren küre şeklinde gövdeleri vardır. En uzun kaktüsün boyu[Not 1] 78 fit (24 m) olarak ölçüldü. Serbest duran kaktüs Pachycereus pringlei 'nin kaydedilen maksimum yüksekliği 19,2 m (63 ft)[5] ve en kısası Blossfeldia liliputiana olgunken sadece yaklaşık 1 cm (0,4 in) dir. Tam büyümüş bir saguaronun (Carnegiea gigantea) bir yağmur fırtınasında 200 ABD galonu (760 L; 170 imp gal) kadar su emebildiği söylenir.[6]
Birkaç türün görünüşü en azından yüzeysel olarak familyanın çoğundan çok farklıdır. Leuenbergeria, Rhodocactus ve Pereskia cinslerinin bitkileri çevrelerinde büyüyen diğer ağaçlara ve çalılara benzer. Kalıcı yaprakları vardır ve daha yaşlı olduklarında kabukla kaplı gövdeleri vardır. Areolleri onları kaktüsler olarak tanımlar ve görünüşlerine rağmen onların da su tasarrufu için birçok uyumu vardır. "Leuenbergeria" tüm kaktüslerin evrimleştiği atalardan kalma türlere yakın kabul edilir.
Tropikal bölgelerdeki diğer kaktüsler ormana tırmanıcı ve epifit (ağaçlarda büyüyen bitkiler) olarak büyür. Sapları genellikle düz, neredeyse yaprak benzeri görünümlü, daha az dikenli veya iyi bilinen Noel kaktüsü veya Şükran Günü kaktüsü (Schlumbergera cinsi) gibi dikensizdir.
Kaktüslerin çeşitli kullanımları vardır: birçok türü süs bitkisi olarak, diğerleri yem veya yiyecek için (özellikle meyveleri) yetiştirilir. Cochineal, bazı kaktüslerde yaşayan ve kırmızı boya yapmakta kullanılan bir böceğin ürünüdür.
Hem Eski hem de Yeni Dünya 'daki sukulentler - bazıları Euphorbiaceae (euphorbias) gibi - kaktüslere çarpıcı benzerlik gösterir ve bu yanlış olabilir ama yaygın kullanımda "kaktüs" olarak adlandırılır.
Çölde yetişen Saguaro en büyük kaktüslerden biridir. Yetişkinleri genellikle 12 metre boya ulaşır, nadiren de 15 metreyi aşanları bulunur.
Kaktüsler çok yağış ve su istemeyen bitkilerdir. Genellikle çöllerde ve sıcak iklimlerde yetişirler. Kökleri çok uzun ve kalındır. Bu özellikleri ve yapraklarının diken şeklinde olması onları diğer bitkilerden ayırır. Kaktüslerin eni ve boyu iyi beslendiği takdirde oldukça uzundur.
Bazı kaktüslerin dikenleri zehirli olabileceği gibi, her şekilde deriye battığında ince dikenleri yüzünden can acıtabilirler ve çıkarılmaları duruma göre zor olabilir. Tüm kaktüs çeşitleri diken bulundurmaz ve peyote gibi dikensiz kaktüs türleri de vardır.
Morfoloji[değiştir | kaynağı değiştir]

1,500 ila 1,800 kaktüs türü çoğunlukla iki "çekirdek kaktüs" grubundan birine girer: opuntias (alt familya Opuntioideae) ve "kaktoidler" (alt familya Cactoideae). Bu iki grubun çoğu üyesi kaktüsler olarak kolayca tanınır. Fotosentez'in ana organları olan etli sukulent gövdeleri vardır. Yaprakları ya yoktur ya da küçük veya geçicidir. Genellikle derinden etli çiçek tablası (çiçek kısımlarının büyüdüğü sap kısmı) içine gömülü çanak yaprakları ve yaprakların altında çiçekler ile tohumlukları bulunur.
Tüm kaktüslerin areole’leri vardır—iğneler, normal sürgünler ve çiçekler üreten kısacık ara düğümleri olan çok özelleşmiş kısa sürgünler.[7]
Kalan kaktüsler üç ağaç benzeri cins, Leuenbergeria, Pereskia ve Rhodocactus (hepsi daha önce Pereskiaya yerleştirilmişti) ve çok daha küçük olan Maihuenia olarak sadece iki gruba ayrılır. Bu iki grup diğer kaktüslerden oldukça farklıdır,[7] ki bu da bir bütün olarak kaktüslerin herhangi bir tanımının sıklıkla onlar için istisnalar yapması gerektiği anlamına gelir. İlk üç cinsin türleri yüzeysel olarak diğer tropik orman ağaçlarına benzer. Olgunlaştıklarında, kabuk ile kaplanabilen odunsu gövdeleri ve fotosentez aracını sağlayan uzun ömürlü yaprakları vardır. Çiçekleri, üst tohumluklara (yani çanak yaprakların ve taç yaprakların bağlanma noktalarının üzerinde) ve daha çok yaprak üreten areollere sahip olabilir.
İki "Maihuenia" türü etli fakat fotosentetik olmayan gövdelere ve belirgin sukulent yapraklara sahiptir.[7]
Büyüme yapıları[değiştir | kaynağı değiştir]
Kaktüsler, açık ve basit kategorilere ayrılması zor olan çok çeşitli büyüme alışkanlıkları gösterir.
- Ağaçsı kaktüsler
Ağaç benzeri (ağaçsı) olabilirler, yani genellikle tepesinde bir ila birçok dalla az ya da çok odunsu gövde vardır. Leuenbergeria, Pereskia ve Rhodocactus cinslerinde dallar yapraklarla kaplı olduğundan bu cinslerin türleri kaktüs olarak tanınmayabilir. Diğer kaktüslerin çoğunda, dallar daha çok kaktüs benzeri, yaprakları ve kabuğu çıplaktır ve Pachycereus pringlei veya daha büyük opuntia'lardaki gibi dikenlerle kaplıdır.
Bazı kaktüsler ağaç boyutunda olabilir ancak daha büyük Echinocactus platyacanthus örnekleri gibi dalları olmayabilir. Kaktüsler ayrıca çalı olarak da tanımlanabilir, Stenocereus thurberi örneği gibi birkaç gövdesi yerden veya çok aşağıdan çıkan dallardan gelir.[8]
- Sütunlu kaktüsler
Daha küçük kaktüsler sütunlu olarak tanımlanabilir. Gövde ve dallara çok net bir bölünme olmaksızın dallanabilen veya ayrılmayan dik, silindir biçimli gövdelerden oluşurlar. Sütunlu formlar ile ağaç benzeri veya çalı formları arasındaki sınırın tanımlanması zordur. Örneğin Cephalocereus senilis türünün giderek daha küçük örnekleri sütunluyken daha yaşlı ve daha büyük örnekler ağaç benzeri olabilir.
Bazı durumlarda, "sütunlar" dikey yerine yatay olabilir. Böylece, Stenocereus eruca toprak boyunca büyüyen, aralıklarla köklenen gövdelere sahiptir.[9]
- Küresel kaktüsler
Sapları daha da küçük olan kaktüsler küresel (veya küre) olarak tanımlanabilir. Sütunlu kaktüslerden daha kısa, daha top şeklindeki gövdelerden oluşurlar. Küresel kaktüsler Ferocactus latispinus gibi yalnız olabilir veya gövdeleri büyük höyükler oluşturabilen kümeler oluşturabilir. Bir kümedeki gövdelerin tümü veya bir kısmı ortak bir kökü paylaşabilir.[9]
- Diğer formlar
Diğer kaktüsler oldukça farklı bir görünüme sahiptir. Tropik bölgelerde, bazıları orman tırmanıcıları ve epifitler olarak yetişir. Sapları genelde yassıdır, görünüşte neredeyse yaprak benzeridir, daha az dikeni vardır veya hiç dikeni yoktur. Tırmanıcı kaktüsleri çok büyük olabilir; bir Hylocereus örneğinin kökten en uzak gövdeye kadar 100 metre (330 ft) uzunluğunda olduğu bildirildi. Rhipsalis veya Schlumbergera türleri gibi epifitik kaktüsler genellikle aşağı doğru sarkar ve yerden yüksek ağaçlarda büyüdükleri yerlerde yoğun kümeler oluşturur.
-
 Ağaç gibi yapı (Pereskia aculeata)
Ağaç gibi yapı (Pereskia aculeata) -
 Ağaç gibi yapı (Pachycereus pringlei)
Ağaç gibi yapı (Pachycereus pringlei) -
 Uzun dalsız sütunlu yapı (Cephalocereus)
Uzun dalsız sütunlu yapı (Cephalocereus) -
 Daha kısa kümelenmiş sütunlu yapı (Ferocactus pilosus)
Daha kısa kümelenmiş sütunlu yapı (Ferocactus pilosus) -
 Yalnız küresel yapı (Ferocactus echidne)
Yalnız küresel yapı (Ferocactus echidne) -
 Kümelenmiş küresel yapı (Rebutia tür)
Kümelenmiş küresel yapı (Rebutia tür) -
 Epifitik kaktüs (Rhipsalis paradoxa)
Epifitik kaktüs (Rhipsalis paradoxa)







Gövdeler[değiştir | kaynağı değiştir]

Yapraksız, dikenli gövde, kaktüslerin çoğunun (ve en büyük alt familya olan Cactoideae'ye ait olanların hepsinin) karakteristik özelliğidir.
Gövde genelde suludur, yani suyu depolamak için uyarlanmıştır. Gövdenin yüzeyi pürüzsüz olabilir (bazı "Opuntia" türlerinde olduğu gibi) veya genellikle tüberkül adı verilen çeşitli türlerde çıkıntılarla kaplı olabilir. Bunlar, Mammillaria cinsindeki küçük "tümsekler"den belirgin, meme ucu benzeri şekillere ve neredeyse Ariocarpus türündeki yapraklara benzeyen çıkıntılara kadar değişir.
Gövde ayrıca kaburgalı veya yivli olabilir. Bu kaburgaların belirginliği, gövdenin ne kadar su depoladığına bağlıdır: dolduğunda (bir kaktüsün kütlesinin %90'ı su olabilir), kaktüs kısa olduğunda kaburgalar şişmiş gövdede neredeyse görünmez olabilir. Su azaldığında ve gövdeler büzüldüğünde kaburgalar çok görünür olabilir.[9]
Çoğu kaktüsün gövdesi yeşilin bir tonudur, genellikle mavimsi veya kahverengimsi yeşildir. Bu tür gövdeler klorofil içerir ve fotosentez yapabilir; ayrıca stoma (gazların geçişine izin vermek için açılıp kapanabilen küçük yapılar) vardır.
Kaktüs gövdeleri genellikle gözle görülür şekilde mumludur.[9]
Areoleler[değiştir | kaynağı değiştir]

.jpg)
.jpg)


Areole'ler kaktüslere özgü yapılardır. Değişken olmakla birlikte, genellikle, dikenlerin çıktığı gövdelerde yünlü veya tüylü alanlar olarak görünürler. Çiçekler de areollerden çıkar. Tüm kaktüslerin atasına benzer olduğuna inanılan Leuenbergeria cinsinde areoller yaprakların aksillerinde (yani yaprak sapı ile gövde arasındaki açıda) bulunur.[10] Yapraksız kaktüslerde, areoller genellikle yaprak tabanlarının olması gereken gövde üzerindeki yükseltilmiş alanlardadır.
Areoleler, son derece özelleşmiş ve çok yoğun sürgünler veya dallardır. Normal bir sürgünde, yaprak veya çiçek taşıyan düğümler gövde uzunluklarıyla (boğum arası) ayrılır. Bir areolde düğümler birbirine çok yakındır, tek bir yapı oluştururlar. Areol dairesel olabilir, oval bir şekle uzatılabilir veya hatta iki parçaya ayrılabilir; iki parça bir şekilde (örneğin gövdedeki bir oluk ile) gözle görülür şekilde bağlantılı olabilir veya tamamen ayrı görünebilir (dimorfik bir areol). Gövdenin tepesine yakın olan kısım daha sonra çiçek diğer kısım ise dikenler üretir. Areoller genellikle, areole tüylü veya yünlü bir görünüm veren, bazen sarı veya kahverengi gibi belirgin bir renkte olan çok hücreli tüylere (trihome'ler sahiptir.[9]
Çoğu kaktüste, areoller sadece birkaç yıllığına yeni dikenler veya çiçekler üretir ve daha sonra etkisizleşir. Bu, nispeten sabit sayıda diken ile sonuçlanır, çiçekler sadece hala büyüyen ve yeni areoller oluşturan gövdelerin uçlarından üretilir. Kaktüslerin atasına yakın bir cins olan “Pereskia”da areoller çok daha uzun süre aktif kalır; bu aynı zamanda Opuntia ve Neoraimondia için de geçerlidir.[9]
Yapraklar[değiştir | kaynağı değiştir]
Kaktüslerin büyük çoğunluğunda görünür yaprak yoktur; fotosentez gövdelerde gerçekleşir (bazı türlerde yassı ve yapraksı olabilir). İstisnalar üç kaktüs grubunda ortaya çıkar. Leuenbergeria, Pereskia ve Rhodocactus türlerinin tüm türleri yüzeysel olarak normal ağaçlar veya çalılar gibidir ve çok sayıda orta damarlı ve her iki tarafta düzleştirilmiş bıçak (lamina) yapraklıdır. Opuntia grubundaki birçok kaktüsün (alt familya Opuntioideae, opuntioidler) ayrıca uzun ömürlü olabilen ('Pereskiopsis türlerinde olduğu gibi) veya sadece büyüme mevsimi boyunca üretilebilen görünen ve sonra kaybolan (birçok Opuntia türünde olduğu gibi) yaprakları vardır.[9] Küçük cins Maihuenia da fotosentez için yapraklara ihtiyaç duyar.[11] Yaprakların yapısı bu gruplar arasında biraz farklıdır. Opuntioidler ve Maihuenia sadece orta damardan oluşuyor gibi görünen yapraklara sahiptir.[12]
Görünür fotosentetik yaprakları olmayan kaktüslerin bile, incelenen türlerin yaklaşık yarısında genellikle 5 mm (0.2 inç) uzunluğundan daha kısa ve neredeyse her zaman 15 milimetre (0,59 in)* uzunluğunda çok küçük yaprakları vardır. Bu tür yaprakların işlevi fotosentez olamaz; auxin gibi bitki hormonlarının üretiminde ve aksiller tomurcukların tanımlanmasında bir rolü olduğu fikri öne sürülmüştür.[13]
Dikenler[değiştir | kaynağı değiştir]
Botanik olarak "iğneler " (ingilizce: spine)" dikenlerden" (İngilizce:"thorn") farklıdır: iğneler değiştirilmiş yapraklar, dikenler ise değiştirilmiş dallardır. Kaktüsler, her zaman yukarıda belirtildiği gibi areollerden dikenler üretir. Pereskia, Pereskiopsis ve Maihuenia gibi yapraklı kaktüslerde bile dikenler vardır, bu yüzden tam yapraksızlıktan önce evrimleştikleri açıktır. Bazı kaktüsler sadece gençken, muhtemelen sadece fideyken dikenlidir. Bu, özellikle Rhipsalis ve Schlumbergera gibi ağaçta yaşayan kaktüsler için değil aynı zamanda Ariocarpus gibi bazı yerde yaşayan kaktüsler için de geçerlidir.[9]
Kaktüslerin dikenleri, türler arasında sayı, renk, boyut, şekil ve sertlik bakımından ve ayrıca bir areol tarafından üretilen tüm dikenlerin benzer olup olmadığı veya farklı türde olup olmadığı konusunda büyük farklılıklar gösterdiğinden, tanımlamada sıklıkla yararlıdır. Çoğu diken düz veya en fazla hafif kavislidir ve uzunluklarına ve kalınlıklarına bağlı olarak saç benzeri, kıl benzeri, iğne benzeri veya bız benzeri olarak tanımlanır. Bazı kaktüslerin düzleştirilmiş dikenleri vardır (örneğin Sclerocactus papyracanthus). Diğer kaktüslerin çengelli dikenleri vardır. Bazen, bir veya daha çok merkezi diken kancalıdır, dış dikenler ise düzdür (örneğin, Mammillaria rekoi).[9]
Normal uzunluktaki dikenlere ek olarak, Opuntioideae alt familyasının üyeleri, uzunlukları boyunca dikenli ve kolayca dökülen glochid adı verilen nispeten kısa dikenlidir. Bunlar cilde girer ve çok ince olmaları ve kolayca kırılmaları nedeniyle çıkarılmaları zordur ve uzun süreli tahrişe neden olur. [9]
-
 Ferocactus’ün dikenleri
Ferocactus’ün dikenleri -
 (cf. Mammillaria rekoi) kancalı merkez dikeni
(cf. Mammillaria rekoi) kancalı merkez dikeni -
 Sclerocactus papyracanthus’un olağanüstü düzleştirilmiş dikenleri
Sclerocactus papyracanthus’un olağanüstü düzleştirilmiş dikenleri -
 Opuntia microdasys’in glokidleri
Opuntia microdasys’in glokidleri




Kökler[değiştir | kaynağı değiştir]
Yerde yaşayan kaktüslerin çoğunda yalnızca yüzeye yakın, bitkinin tabanı etrafında değişen mesafelerde yayılan ince kökler bulunur. Ariocarpus gibi bazı kaktüs cinslerinde kazık kök'ler vardır, bunlar vücuttan önemli ölçüde daha büyük ve daha büyük hacimlidir. Kazık kökler, daha büyük sütunlu kaktüslerin dengelenmesine yardımcı olabilir.[14] Tırmanan, sürünen ve epifitik kaktüsler, yalnızca köklenme ortamıyla temas ettikleri gövdeler boyunca üretilen adventif kökler'e sahip olabilir.[9]
Çiçekler[değiştir | kaynağı değiştir]



Dikenleri gibi kaktüsler de çiçek değişkendir. Genellikle, tohumluk, gövdeden veya hazne dokusundan türetilen malzeme ile çevrilidir ve perikarpel adı verilen bir yapı oluşturur. Yapraklardan ve sepallerden türetilen doku, perikarpeli devam ettirerek kompozit bir tüp oluşturur- tamamına çiçek tüpü denebilir ancak kesinlikle sadece tabandan en uzak kısım çiçek kökenlidir. Boru şeklindeki yapının dışında genellikle yün ve diken üreten areoller bulunur. Genellikle, tüp ayrıca, yavaş yavaş sepal benzeri ve daha sonra petal benzeri yapılara dönüşen küçük ölçek benzeri braktelere sahiptir, bu nedenle sepaller ve petaller açıkça ayırt edilemez (ve bu nedenle genellikle "tepaller" denilir).[9] Bazı kaktüsler, yün veya dikensiz çiçek tüpleri üretir (örneğin Gymnocalycium)[15] veya herhangi bir dış yapıdan tamamen yoksundur (örneğin Mammillaria).[9]
Diğer kaktüslerin çoğunun çiçeklerinden farklı olarak Pereskia çiçekleri salkımlarda doğabilir.[10]
Kaktüs çiçeklerinde genellikle birçok erkek organ ancak yalnızca tek dişi organ vardır, bu da sonunda birden fazla tepecik (ingilizce: stigma)olarak dallanabilir. Ercikler genellikle çiçek tüpünün üst kısmının tüm iç yüzeyinden çıkar, ancak bazı kaktüslerde organlarındakiler çiçek tüpünün iç kısmının daha özel alanlarında bir veya daha fazla farklı "seri" olarak üretilir.[9]
Bir bütün olarak çiçek genellikle radyal olarak simetriktir (aktinomorfik), ancak bazı türlerde iki taraflı simetrik olabilir (zigomorfik). Çiçek renkleri beyazdan sarıya ve kırmızıdan morumsu kırmızı renge kadar değişir.[9]
Su tasarrufu için uyum sağlama[değiştir | kaynağı değiştir]
Tüm kaktüslerin verimli su kullanımı için ortama bazı uyum sağlama düzenleri vardır. Kaktüslerin çoğu—opuntialar ve cactoidler—sıcak ve kuru ortamlarda (örn. Kserofitler) yaşayabilir ama modern kaktüslerin ilk ataları zaten zaman zaman rastlanan kuraklık dönemlerine uyum sağlamışlardı.[7] Hylocereeae ve Rhipsalideae oymaklarındaki birkaç kaktüs türü su tasarrufunun önemli olmadığı tropik ormanlarda tırmanıcılar veya epifitler olarak yaşama uyum sağlamıştı.
Yapraklar ve dikenler[değiştir | kaynağı değiştir]
Görünür yaprakların olmaması çoğu kaktüsün en çarpıcı özelliklerinden biridir. Pereskia (bütün kaktüslerin evrimleştiği ata türlerine yakın olan) uzun ömürlü yapraklıdır ancak birçok türlerde kalınlaşmış ve sukulentir.[7] Opuntioid Pereskiopsis gibi uzun ömürlü yaprakları olan diğer kaktüs türlerinin de etli yaprakları vardır.[16] Su tutmada önemli olan yüzey alanının hacme oranıdır. Su kaybı yüzey alanıyla orantılıyken mevcut su miktarı ise hacimle orantılıdır. İnce yapraklar gibi yüzey alanı-hacim oranı fazla yapılar, kalınlaşmış gövdeler gibi alan-hacim oranı az olan yapılardan zorunlu olarak daha çok su kaybeder.
Yaprakların değişmiş hali olan dikenler, gerçek yapraklı kaktüslerde bile bulunur ve dikenlerin yaprak kaybından önceki evrimini gösterir. Dikenlerin yüzey alanı-hacim oranı yüksek olmasına rağmen, olgunlukta çok az su içerirler veya hiç su içermezler ve ölü hücrelerden oluşan liflerden oluşurlar.[12] Dikenler bazı türlerde otoburlara karşı koruma ve kamuflaj sağlar ve çeşitli şekillerde suyun korunmasına yardımcı olur. Kaktüsün yüzeyine yakın havayı hapseder, buharlaşmayı ve terleme’yi azaltan daha nemli bir tabaka oluştururlar. Kaktüsün yüzey sıcaklığını ve aynı zamanda su kaybını azaltan bir miktar gölge de yapabilirler. Sis veya sabahın erken saatlerinde sis sırasında olduğu gibi yeterince nemli hava mevcut olduğunda, dikenler su buharını yoğunlaştırabilir, bu ise daha sonra yere damlar ve kökler tarafından emilir.[9]
Gövdeler[değiştir | kaynağı değiştir]

Kaktüslerin çoğu gövde suludur (İngilizce: stem succulent) yani gövdenin su depolamak için kullanılan ana organ olduğu bitkilerdir. Bir kaktüsün toplam kütlesinin %90'ını su oluşturabilir. Gövde şekilleri kaktüsler arasında çok farklılık gösterir. Sütunlu kaktüslerin silindirik şekli ve küresel kaktüslerin küresel şekli, düşük yüzey alanı-hacim oranı oluşturur, böylece su kaybını azaltır ve güneş ışığının ısıtma etkilerini en aza indirir. Pek çok kaktüsün yivli veya oluklu gövdeleri, kuraklık dönemlerinde gövdenin büzülmesine ve daha sonra müsaitlik dönemlerinde suyla dolduğunda şişmesine izin verir.[9] Olgun bir saguaro'nun (Carnegiea gigantea) bir yağmur fırtınası sırasında 200 ABD galonu (760 L; 170 imp gal) kadar su emebildiği söylenir.[6] Gövdenin dış tabakası genellikle su kaybını azaltan mumsu tabakalarla güçlendirilmiş sert bir kütikülaya sahiptir. Bu katmanlar, birçok kaktüsün gövde rengindeki grimsi veya mavimsi renkleri yapar.[9]
Çoğu kaktüsün gövdesinde yaprakların yokluğunda fotosentez yapmalarına imkan veren uyarlamaları vardır. Bu konu, aşağıda Metabolizma altında daha ayrıntılı olarak işlenmektedir.
Kökler[değiştir | kaynağı değiştir]
Birçok kaktüsün geniş bir alana yayılmış kökleri olur, ancak toprağa yalnızca kısa bir mesafe nüfuz ederler. Bir keresinde incelenen 2 m (7 ft) boyundaki genç bir saguaro’nun yalnızca 12 cm (4,7 in) çapında bir kök sistemi vardı ama kökleri en çok 10 cm (4 in) derinlikteydi.[14] Kaktüsler ayrıca kuraklıktan sonra yağmur yağdığında hızla yeni kökler oluşturabilir. Kaktüslerin kök hücrelerindeki tuz konsantrasyonu nispeten yüksektir.[17] Tüm bu uyumlar, kaktüslerin kısa veya hafif yağış dönemlerinde suyu hızla emmesini sağlar. Bu nedenle bildirildiğine göre, Ferocactus cylindraceus 7 mm (0,3 in) kadar az yağmur yağdığında bile yağıştan 12 saat içinde çok miktarda su alabilir ve birkaç günde tam sulu olur.[9]
Çoğu kaktüste gövde suyu depolamak için ana organ görevi görse de, bazı kaktüslerin ek olarak büyük kazık kökleri (İngilizce: Taproot) vardır.[9] Bunlar, dünyanın en kurak yerlerinden birinde kuzey Şili'deki Atacama Çölü’nde yetişen Copiapoa atacamensis,[9] gibi türler söz konusu olduğunda kökler yer üstü gövdesinin uzunluğunun birkaç katı olabilir. [18]


Metabolizma[değiştir | kaynağı değiştir]
Fotosentez, bitkilerin karbon dioksit gazını (CO2) almasını gerektirir. Bunu yaparken terleme yoluyla su kaybederler. Diğer sukulentler gibi, kaktüsler de fotosentez yaparak bu su kaybını azaltır. "Normal" yapraklı bitkilerC3 mekanizmasını kullanır: gündüz saatlerinde, CO2 sürekli olarak yaprakların içindeki boşluklarda bulunan havadan çekilir ve önce dönüştürülür üç karbon atomu içeren bir bileşiğe (3-fosfogliserat) ve daha sonra karbonhidrat gibi ürünlere dönüştürülür. Bitki içindeki havanın iç boşluklara erişimi, açılıp kapanabilen stomata tarafından kontrol edilir. Fotosentez sırasında sürekli bir CO2 kaynağına duyulan ihtiyaç, stomaların açık olması gerektiği anlamına gelir, bu nedenle su buharı sürekli olarak kaybolur. C3 mekanizmasını kullanan bitkiler kökleri yoluyla aldıkları suyun %97'sini bu şekilde kaybederler.[19] Bir başka sorun da sıcaklık arttıkça, CO2 yakalayan enzim bunun yerine giderek daha fazla oksijen yakalamaya başlar ve fotosentez verimliliğini %25'e kadar azaltır.[20]
Taksonomi ve sınıflandırma[değiştir | kaynağı değiştir]


(aşağıda) Melocactus, muhtemelen Avrupalılar tarafından görülen ilk cinstir
Bilim için ilk kaktüslerin keşfedilmesinden bu yana kaktüsleri adlandırmak ve sınıflandırmak hem zor hem de tartışmalı olmuştur. Zorluklar Carl Linnaeus ile başladı. 1737'de tanıdığı kaktüsleri "Cactus" ve "Pereskia" olmak üzere iki cinse ayırdı. Bununla birlikte, modern botanik terminolojinin başlangıç noktası olan 1753'te Species Plantarum yayınladığında hepsini tek bir cinse, Cactuse indirdi. "Kaktüs" kelimesi, Theophrastus tarafından muhtemelen yabani enginar (Cynara cardunculus) dikenli bitkisinin ismi için kullanılan Antik Yunanca Grekçe: κάκτος (kaktos) kelimesinden Latince türetilmiştir.[21][22]
1754'te Philip Miller gibi sonraki botanikçiler, kaktüsleri birkaç cinse ayırdı ve 1789'da Antoine Laurent de Jussieu yeni yarattığı Cactaceae familyasına yerleştirdi. 20. yüzyılın başlarında botanikçiler, Linnaeus'un “Cactus” adının anlamı konusunda (cins mi yoksa familya mı ?) o kadar karıştırıldığını ve bir cins adı olarak kullanılmaması gerektiğini hissetmeye başladılar. 1905 Viyana botanik kongresi "Cactus" adını reddetmiş ve bunun yerine "Mammillaria"nın Cactaceae familyasının tür cinsi olduğunu ilan etmiştir. Ancak, Cactaceae adını korunan ad yaptı ve Cactaceae familyasının artık adını aldığı cinsi içermediği olağandışı duruma yol açtı.[23]
Zorluklar, kısmen bitkilere bilimsel isimler vermenin "tip örnekler"e dayanması nedeniyle devam etti. Nihayetinde, eğer botanikçiler belirli bir bitkinin örneğin "Mammillaria mammillaris"in bir örneği olup olmadığını bilmek istiyorlarsa, onu bu ismin kalıcı olarak iliştirildiği tür örneğiyle karşılaştırabilmeleri gerekir. Tip numuneleri normalde sıkıştırma ve kurutma ile hazırlanır, ardından kesin referanslar olarak işlev görmesi için herbaria içinde saklanır. Ancak kaktüsleri bu şekilde muhafaza etmek çok zordur; kurumaya direnecek şekilde evrimleşmişlerdir ve gövdeleri kolayca sıkışmaz.[24]
Diğer bir zorluk ise, birçok kaktüse botanikçiler yerine yetiştiriciler ve bahçıvanlar tarafından isim verilmiş olmasıdır; sonuç olarak, Algler, mantarlar ve bitkiler için Uluslararası İsimlendirme Kodu (kaktüslerin ve diğer bitkilerin adlarını düzenleyen) hükümleri genellikle göz ardı edildi. Curt Backeberg'in, özellikle, David Hunt'a göre, bir örneğe hiçbir ismi eklenmeden 1,200 türü adlandırdığı veya yeniden adlandırdığı söylenir. "Muhtemelen yüzyıllar boyunca kaktüs taksonomistlerini rahatsız edecek bir isimlendirme kaos izi bıraktı."[25]
Sınıflandırma[değiştir | kaynağı değiştir]
1984 yılında, Uluslararası Sukulent Bitki Çalışma Organizasyonu'nun (İngilizce: International Organization for Succulent Plant Study) Cactaceae Bölümünün cinsler düzeyinde fikir birliği sınıflandırmaları yapmak için Uluslararası Cactaceae Sistematik Grubu (ICSG) (İngilizce: the International Cactaceae Systematics Group) adlı çalışma grubu kurması gerektiğine karar verildi. Sistemleri, sonraki sınıflandırmaların temeli olarak kullanıldı. 21. yüzyılda yayınlanan ayrıntılı muameleler, familya yaklaşık 125-130 cins ve 1,400-1,500 türe ayrıldı ve bunlar daha sonra bir dizi kabile ve alt familyaya ayrıldı.[26][27][28] Kaktüs familyasının ICSG sınıflandırması, en büyüğü dokuz kabileye bölünmüş dört alt familyayı tanıdı. Alt familyalar şunlardı:[26]
- Pereskioideae alt familyası K. Schumann
- ICSG klasmanındaki tek cins Pereskia idi. Cactaceae'nin atalarına en yakın olduğu düşünülen özelliklere sahiptir. Bitkiler, yapraklı ağaçlar veya çalılardır; gövdeleri, yivli veya tüberküllü olmaktan ziyade kesitte düzgün bir şekilde yuvarlaktır.[26] fotosentez'de iki sistem kullanılabilir, her ikisi de "normal" C3 karbon tutulumu mekanizması ve Krassulasean asit metabolizması (CAM)—kaktüslerin ve sukulentlerin "gelişmiş" özelliğini yani su tasarrufunu sağlar.[7]Moleküler filogenetik çalışmaları, geniş anlamda sınırlı olduğunda, Pereskianın monofiletik olmadığını ve[28][29] bölündüğünü gösterdi. Üç cins, Leuenbergeria, Rhodocactus ve dar bir şekilde sınırlandırılmış Pereskia.[30][31] "Leuenbergeria" daha sonra kendi başına ayrı bir monojenerik alt familya olan Leuenbergerioideae'ye yerleştirilir.[32]
- Alt familya Opuntioideae K. Schumann
- Bu alt familyaya yaklaşık 15 cins dahildir. Gençken yaprakları olabilir ancak bunlar daha sonra kaybolur. Sapları genellikle farklı "eklemlere" veya "pedlere" (cladodelara) ayrılır.[26]
Bitkilerin boyutları, Maihueniopsis[33] küçük yastıklarından, 10 m (33 ft)'e ya da daha fazlaya yükselen ağaç benzeri Opuntia türlerine kadar değişir [34]
- Tek cins “Maihuenia” olup, her ikisi de alçakta büyüyen hasır yapan iki türdür. Kaktüsler içinde ilkel olan bazı özelliklere sahiptir. Bitkilerin yaprakları vardır ve crassulacean asit metabolizması tamamen yoktur.[26]
- Alt familya Cactoideae
- Dokuz kabileye bölünmüş, bu, tüm "tipik" kaktüsler dahil en büyük alt ailedir. Üyeler, ağaç benzerinden epifitike kadar değişen alışkanlıklarda oldukça değişkendir. Yapraklar normalde yoktur, ancak bazen genç bitkiler tarafından çok azaltılmış yapraklar üretilir. Saplar genellikle parçalara ayrılmamıştır ve nervürlü veya tüberküllüdür. Kabilelerden ikisi, Hylocereeae ve Rhipsalideae, oldukça farklı bir görünüme sahip tırmanma veya epifitik formları içerir; sapları düzleştirilmiştir ve parçalara ayrılabilir.[26]
Moleküler filogenetik çalışmalar bu alt familyaların üçünün (Pereskioideae değil) monofilisini desteklemiştir,[28][29] ancak bu seviyenin altındaki tüm kabileleri ve hatta cinsleri desteklememiştir; gerçekten de 2011'de yapılan bir araştırma, araştırmada örneklenen Cactoideae alt familyasındaki cinslerin yalnızca %39'unun monofilik olduğunu buldu.[28] Kaktüslerin sınıflandırılması şu anda belirsizliğini korur ve muhtemelen değişecektir.
Bakımı[değiştir | kaynağı değiştir]
Kaktüse yılda en az bir kere vitamin ve mineral desteği sağlanmalı, toprağı tazelenip havalandırılmalıdır. Saksı değişimi 5-6 yılda bir kere yeterlidir. Seyrek aralıklarla özellikle yazın az su damlatılabilir.
Yetiştirme[değiştir | kaynağı değiştir]

Kaktüslerin popülaritesi, birçok kitabın kaktüs yetiştirilmesi için yazıldığından anlaşılabilir. Kaktüsler doğal olarak çok çeşitli habitatlarda bulunur ve farklı iklimlere sahip birçok ülkede yetiştirilir, bu nedenle bir türün normal olarak büyüdüğü koşulları tam olarak kopyalamak genellikle pratik değildir.[35] Farklı koşullara ihtiyaç duyan ve en iyi ayrı ayrı yetiştirilen yarı çöl kaktüsleri ve epifitik kaktüsler arasında geniş bir ayrım yapılabilir.[36] Bu bölüm esasen buna imkan veren iklimlerde dışarıda toprakta yetiştirmekten ziyade, bir serada veya evde olduğu gibi kaplarda ve koruma altında yarı çöl kaktüslerinin yetiştirilmesiyle ilgilidir. Epifitik kaktüslerin yetiştirilmesi için, bkz. Schlumbergera' Yetiştiriciliği (Noel veya Şükran Günü kaktüsleri) ve Epiphyllum melezlerinin yetiştirilmesi.
Yetiştirme ortamı[değiştir | kaynağı değiştir]

Yetiştirme ortamının amacı bitkiyi beslemek için destek sağlamak, su, oksijen ve çözünmüş mineralleri depolamaktır.[37] Kaktüsler söz konusu olduğunda, çok havadar açık bir ortamın önemli olduğu konusunda genel bir fikir birliği vardır. Kaktüsler saksılarda yetiştirildiğinde, bunun nasıl sağlanacağına dair öneriler büyük ölçüde değişir; Miles Anderson, mükemmel bir yetiştirme ortamı tanımlaması istendiğinde, "on yetiştiricinin 20 farklı cevap vereceğini" söyler.[38] Roger Brown, solucan gübrelerinden toprak ilavesiyle iki kısım ticari topraksız büyüme ortamı, bir kısım hidroponik kil ve bir kısım kaba pomza veya perlit karışımını önerir.[37] Genellikle %25-75 oranında organik esaslı malzeme, geri kalana pomza, perlit veya kum gibi inorganik tavsiyesi diğer kaynaklar tarafından desteklenir. [38][39][40][41] Ancak, organik materyalin kullanımı başkaları tarafından tamamen reddedilir; Hecht, kaktüslerin (epifitler dışında) "humus içeriği düşük veya içermeyen toprak istediğini" söyler ve yetiştirme ortamının temeli olarak kaba kumu tavsiye eder.[42]
Sulama[değiştir | kaynağı değiştir]
Yarı çöl kaktüsleri dikkatli sulamaya ihtiyaç duyar. Genel tavsiye vermek zordur, çünkü gerekli sulama sıklığı kaktüslerin nerede yetiştirildiğine, yetiştirme ortamının doğasına ve kaktüslerin orijinal habitatına bağlı olarak değişir.[43] Brown, "zamansız su verilmesi nedeniyle başka herhangi bir nedenle olduğundan daha fazla kaktüsün öldüğünü" ve uykuda kış mevsiminde bile kaktüslerin biraz suya ihtiyacı olduğunu söyler.[44] Diğer kaynaklar, kışın (Kuzey Yarımküre'de Kasım-Mart arası) suyun verilmeyeceğini söyler.[36] Bir diğer konu ise suyun sertliğidir ki sert su kullanılması gerektiğinde, tuz oluşumunu önlemek için düzenli yeniden saksılama önerilir.[44] Verilen genel tavsiye, büyüme mevsimi boyunca kaktüslerin kapsamlı sulamalar arasında kurumasına izin verilmesi gerektiğidir.[44][45][36] Bir su miktarı ölçer (İngilizce: water meter), toprağın ne zaman kuru olduğunu belirlemede yardımcı olabilir.[45]
Işık ve sıcaklık[değiştir | kaynağı değiştir]
Yarı çöl kaktüsleri vahşi doğada yüksek ışık seviyelerine maruz kalabilse de, yaz aylarında bir seranın daha yüksek ışık seviyelerine ve sıcaklıklarına maruz kaldıklarında yine de biraz gölgelendirmeye ihtiyaç duyabilirler.[46][47] Sıcaklığın 32 °C (90 °F) üzerine çıkmasına izin verilmesi önerilmez.[47] Gereken minimum kış sıcaklığı ilgili kaktüs türlerine çok bağlıdır. Karma bir koleksiyon için Melocactus ve Discocactus gibi soğuğa duyarlı türler hariç olmak üzere genellikle 5 °C (41 °F) ile 10 °C (50 °F) arasında minimum bir sıcaklık önerilir.[48][36] Bazı kaktüsler, özellikle yüksek And dağlarının kaktüsleri, kuru tutulduklarında tamamen dona dayanıklıdır (örneğin Rebutia minuscula, yetiştirirken −9 F sıcaklığa kadar dayanır[49]) ve soğuk bir döneme maruz kaldığında daha iyi çiçek açabilir.[50]
Çoğaltma[değiştir | kaynağı değiştir]
Kaktüsler tohumla, çelikle veya aşılama ile çoğaltılabilir. Yılın başlarında ekilen tohumlar, daha uzun bir büyüme döneminden yararlanan fideler üretir.[51] Tohum nemli yetiştirme ortamına ekilir ve sonra kurumasını önlemek için çimlenmeden 7-10 gün sonraya kadar kapalı bir ortamda tutulur.[52] Çok ıslak yetiştirme ortamı hem tohumların hem de fidelerin çürümesine neden olabilir.[53] Çimlenme için 18-30 °C (64-86 °F) sıcaklık aralığı önerilir; 22 °C (72 °F) civarındaki toprak sıcaklıkları en iyi kök gelişimini destekler. Çimlenme sırasında düşük ışık seviyeleri yeterlidir, ancak daha sonra yarı çöl kaktüsleri güçlü büyüme sağlamak için daha yüksek ışık seviyelerine ihtiyaç duyar ancak daha yüksek sıcaklıklar ve güçlü güneş ışığı gibi bir seradaki koşullar için iklimlendirme gerekir.[52]

Kesimlerle çoğaltma, bitkinin kök salabilen kısımlarını kullanır. Bazı kaktüsler, ayrılabilen veya temiz bir şekilde kesilebilen "pedler" veya "eklemler" üretir. Diğer kaktüsler, çıkarılabilen çıkıntılar üretir.[51] Aksi takdirde, ideal olarak nispeten yeni büyümeden gövde kesimleri yapılabilir. Kesilen yüzeyin üzerinde bir kallus kültürü oluşana kadar herhangi kesilen yüzeyin birkaç gün ila birkaç hafta arasında kurumasına izin verilmesi önerilir. Köklenme daha sonra uygun bir yetiştirme ortamında 22 °C (72 °F) civarında bir sıcaklıkta gerçekleşebilir.[51][52]
Yetiştiricilikte iyi büyümesi zor veya bağımsız büyüyemeyen türler örneğin beyaz, sarı veya kırmızı gövdeli bazı klorofil ‘siz formlar için veya anormal büyüme gösteren bazı formlar için (örneğin, kristat veya monstrose formları) aşılama kullanılır. Yetiştirici, ev sahibi (anaç) bitki için ekimde güçlü büyüyen ve çoğaltılacak bitkiyle uyumlusunu seçer: aşılık filiz. Yetiştirici, hem stok hem de aşılık filiz üzerinde kesintiler yapıp ikisini birleştirir, birleşirken onları birbirine bağlar. Hem kalem hem de stokun benzer çaplarda olduğu düz aşılar ve stokta yapılmış bir yarığa daha küçük bir kalemin yerleştirildiği yarık aşılar gibi çeşitli aşı türleri kullanılır.[54]
Ticari olarak her yıl çok sayıda kaktüs üretilir. Örneğin, 2002'de yalnızca Kore'de, neredeyse 9 milyon ABD Doları değerinde 49 milyon bitki çoğaltıldı. Çoğu (31 milyon bitki) aşılama ile çoğaltılmıştır.[55]
Zararlılar ve hastalıklar[değiştir | kaynağı değiştir]
Bir dizi haşere ekim sırasında kaktüslere saldırır. Bitki özsuyuyla beslenenler arasında hem gövde hem de kök üzerinde yaşayan unlu böceği (ingilizce:mealybug); genellikle sadece gövdelerde bulunan ölçekli böcekler (ingilizce:scale insect); kaktüslerin "nadiren" zararlısı olduğu söylenen beyazsinekler;[56] çok küçük ancak çok sayıda olabilen, etraflarında ince bir ağ oluşturup kaktüsü öldürmeseler bile özsuyunu emerek kötü bir şekilde kaktüsü bozabilen kırmızı örümcek akarları; ve özellikle çiçeklere saldıran thrip’ler. Mevcut biyolojik kontroller olmasına rağmen, bu zararlılardan bazıları birçok böcek ilacına dirençlidir. Kaktüslerin kökleri sciarid sinekler ve mantar sivrisineklerinin larvaları tarafından yenebilir. Sümüklüböcekler ve salyangozlar da kaktüs yerler.[57][58]
Mantarlar, bakteriler ve virüsler, özellikle bitkiler aşırı sulandığında ilk ikisi olan kaktüslere saldırır. Fusarium rot bir yaradan girebilir ve kırmızı-mor küfün eşlik ettiği çürümeye neden olabilir. "Helminosporium çürümesinin nedeni Bipolaris cactivora (syn. Helminosporium cactivorum[59]); Phytophthora türleri de kaktüslerde benzer çürümelere neden olur. Mantar öldürücüler bu hastalıklarla mücadelede sınırlı etkilidir.[60] Kaktüslerde kaktüs virüsü X dahil olmak üzere birçok virüs bulunmuştur. Bunlar, klorotik (soluk yeşil) lekeler ve mozaik etkiler (çizgiler ve daha soluk renkli lekeler) gibi yalnızca sınırlı görünür işaretlere neden olur gibi görünürler.[61] Bununla birlikte, bir "Agave" türünde, kaktüs virüsü X'in, özellikle kökler kuru olduğunda büyümeyi azalttığı gösterilmiştir.[62]
Virüs hastalıklarının tedavisi yoktur.[60]
Galeri[değiştir | kaynağı değiştir]
-
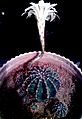 Kaktüs çiçeği
Kaktüs çiçeği -
 Kaktüs çiçeği
Kaktüs çiçeği -
 Kaktüs çiçeği
Kaktüs çiçeği

.jpg)
.jpg)
Notlar[değiştir | kaynağı değiştir]
- ^ yaşayan kaktüs, "Pachycereus pringlei" türünün bir örneğidir. Ölçülen en uzun kaktüs Temmuz 1986'da fırtınada devrilen kolsuz saguaro tipi bir kaktüstü; boyu 78 fit (24 m) idi."Windstorm Fells 78-Foot Cactus--Tallest in World". 29 Ekim 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 4 Ağustos 2015.
Kaynakça[değiştir | kaynağı değiştir]
- ^ Angiosperm Phylogeny Group (2009). "An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III". Botanical Journal of the Linnean Society. 161 (2): 105-121. doi:10.1111/j.1095-8339.2009.00996.x.
- ^ Angiosperm Phylogeny Group (2016). "An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV" (PDF). Botanical Journal of the Linnean Society. 181 (1): 1–20. doi:10.1111/boj.12385. ISSN 0024-4074.
- ^ Christenhusz, M. J. M.; Byng, J. W. (2016). "The number of known plants species in the world and its annual increase". Phytotaxa. 261 (3): 201-217. doi:10.11646/phytotaxa.261.3.1. 29 Temmuz 2016 tarihinde kaynağından arşivlendi.
- ^ Johnson, A.T.; Smith, H.A.; Stockdale, A.P. (2019), Plant Names Simplified : Their Pronunciation Derivation & Meaning, Sheffield, Yorkshire: 5M Publishing, ISBN 9781910455067, p. 26
- ^ Salak, M. (2000), "In search of the tallest cactus", Cactus and Succulent Journal, 72 (3)
- ^ a b Views of the National Parks: Stop #3 - Saguaro (Carnegiea gigantea), National Park Service, US Department of the Interior, 26 Ekim 2011 tarihinde kaynağından arşivlendi, erişim tarihi: 19 Şubat 2012
- ^ a b c d e f Edwards, E.J.; Donoghue, M.J. (2006), "Pereskia and the origin of the cactus life-form" (PDF), The American Naturalist, 167 (6), ss. 777-793, doi:10.1086/504605, PMID 16649155, 13 Şubat 2012 tarihinde kaynağından (PDF) arşivlendi, erişim tarihi: 8 Şubat 2012
- ^ Anderson (2001)
- ^ a b c d e f g h i j k l m n o p q r s t u Anderson (2001), ss. 15–37
- ^ a b Anderson (2001), s. 566
- ^ a b Anderson (2001), s. 398
- ^ a b Mauseth (2007), s. 845
- ^ Mauseth, James D. (2007), "Tiny but complex foliage leaves cccur in many 'leafless' cacti (Cactaceae)", International Journal of Plant Sciences, 168 (6), ss. 845-853, doi:10.1086/518273, p. 845
- ^ a b Biology of Cacti, Dalhousie University, 20 Şubat 2012 tarihinde kaynağından arşivlendi, erişim tarihi: 13 Şubat 2012
- ^ Anderson (2001), ss. 347–348
- ^ Anderson (2001), s. 572
- ^ Kaynak hatası: Geçersiz
<ref>etiketi;GibsNobe90isimli refler için metin sağlanmadı (Bkz: Kaynak gösterme) - ^ Anderson (2001), s. 174
- ^ Kaynak hatası: Geçersiz
<ref>etiketi;RaveEdwa01isimli refler için metin sağlanmadı (Bkz: Kaynak gösterme) - ^ Kaynak hatası: Geçersiz
<ref>etiketi;Shar88isimli refler için metin sağlanmadı (Bkz: Kaynak gösterme) - ^ Johnson, A.T.; Smith, H.A. (1972), Plant Names Simplified : Their Pronunciation Derivation & Meaning, Buckenhill, Herefordshire: Landsmans Bookshop, ISBN 978-0-900513-04-6, p. 19
- ^ Sonnante, G.; Pignone, D.; Hammer, K (2007), "The Domestication of Artichoke and Cardoon: From Roman Times to the Genomic Age", Annals of Botany, 100 (5), ss. 1095-1100, doi:10.1093/aob/mcm127, PMC 2759203 $2, PMID 17611191
- ^ Anderson (2001), s. 96
- ^ Anderson (2001), ss. 93–94
- ^ Anderson (2001), s. 98
- ^ a b c d e f Anderson (2001), ss. 99–103
- ^ Kaynak hatası: Geçersiz
<ref>etiketi;Hunt06isimli refler için metin sağlanmadı (Bkz: Kaynak gösterme) - ^ a b c d Kaynak hatası: Geçersiz
<ref>etiketi;BarcYessHawk11isimli refler için metin sağlanmadı (Bkz: Kaynak gösterme) - ^ a b Kaynak hatası: Geçersiz
<ref>etiketi;EdwaNyffDono05isimli refler için metin sağlanmadı (Bkz: Kaynak gösterme) - ^ Kaynak hatası: Geçersiz
<ref>etiketi;Lode13isimli refler için metin sağlanmadı (Bkz: Kaynak gösterme) - ^ Kaynak hatası: Geçersiz
<ref>etiketi;AsaiMiya16isimli refler için metin sağlanmadı (Bkz: Kaynak gösterme) - ^ Kaynak hatası: Geçersiz
<ref>etiketi;MaytMoli15isimli refler için metin sağlanmadı (Bkz: Kaynak gösterme) - ^ Anderson (2001), s. 399
- ^ Anderson (2001), s. 485
- ^ Keen (1990), s. 15
- ^ a b c d Innes (1995), s. 22
- ^ a b Brown (2001), s. 87
- ^ a b Anderson (1999), s. 217
- ^ Hewitt (1993), s. 147
- ^ Innes (1995), s. 23
- ^ Keen (1990), ss. 27–28
- ^ Hecht (1994), s. 140
- ^ Pilbeam (1987), s. 10
- ^ a b c Brown (2001), s. 88
- ^ a b Hewitt (1993), s. 151
- ^ Brown (2001), s. 85
- ^ a b Hewitt (1993), s. 150
- ^ Pilbeam (1987), s. 11
- ^ Amos, Robert (2012), "Show Reports: Malvern Show", The Alpine Gardener, 80 (1), ss. 80-83
- ^ Sheader, Martin (2012), "Show Reports: Summer Show South", The Alpine Gardener, 80 (1), ss. 88-91
- ^ a b c Innes (1995), s. 28
- ^ a b c Brown (2001), s. 92
- ^ Innes (1995), s. 27
- ^ Innes (1995), s. 29
- ^ Jeong, Myeong Il; Cho, Chang-Hui; Lee, Jung-Myung (2009), Production and Breeding of Cacti for Grafting in Korea, Gyeonggi-do Agricultural Research & Extension Services, 28 Mayıs 2013 tarihinde kaynağından arşivlendi, erişim tarihi: 28 Mart 2012
- ^ Innes (1995), s. 32
- ^ Innes (1995), ss. 31–32
- ^ Brown (2001), ss. 90–91
- ^ "Bipolaris cactivora (Petr.) Alcorn", Species Fungorum, 14 Mayıs 2013 tarihinde kaynağından arşivlendi, erişim tarihi: 30 Mart 2012
- ^ a b Hecht (1994), s. 152
- ^ Duarte, L.M.L.; Alexandre, M.A.V.; Rivas, E.B.; Harakava, R.; Galleti, S.R.; Barradas, M.M. (2008), "Potexvirus diversity in Cactaceae from São Paulo State in Brazil", Journal of Plant Pathology, 90 (3), ss. 545-551, 14 Eylül 2012 tarihinde kaynağından arşivlendi, erişim tarihi: 30 Mart 2012
- ^ Izaguirre-Mayoral, Maria Luisa; Marys, Edgloris; Olivares, Elizabeth; Oropeza, Tamara (1995), "Effect of seasonal drought and cactus X virus infection on the crassulacean acid metabolism of Agave sisalana plants growing in a neotropical savanna", Journal of Experimental Botany, 46 (6), ss. 639-646, doi:10.1093/jxb/46.6.639